
۱۳
بهمن

قابل توجه دانش آموزان پایه هفتم جهت دانلود و استفاده از محتوی کتاب علوم تکمیلی (ویژه دانش آموزان مراکز استعدادهای درخشان) می توانید فایل PDF این کتاب را از طریق لینک زیر دانلود نماییید.
با توجه به اینکه حجم فایل کتاب علوم تکمیلی حدود 16 مگا بایت می باشد جهت سهولت در دانلود این فایل در 3 قسمت آپلود می شود
 دانلود قسمت اول(فهرست تا فصل4) دانلود قسمت دوم (فصل 5 تا 10 )
دانلود قسمت اول(فهرست تا فصل4) دانلود قسمت دوم (فصل 5 تا 10 )دانلود قسمت سوم (فصل 11 تا 15)
۰۷
بهمن
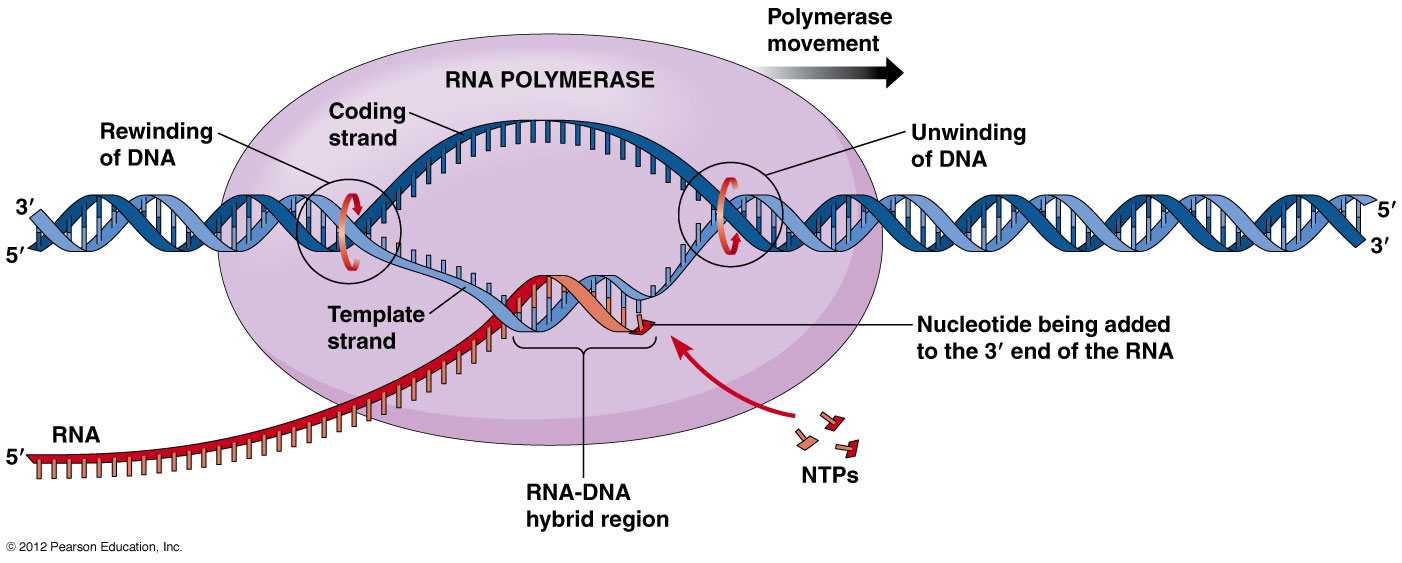
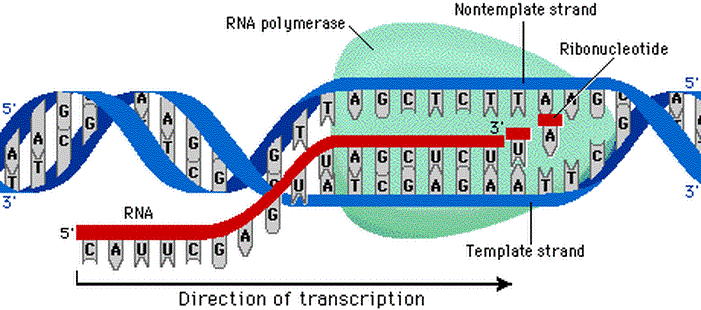
در هسته هر سلول مولکولهایی قرار دارند که اساسی ترین اطلاعات حیات را در خود ذخیره کرده اند. این مولکول ها، دئوکسی ریبو نوکلئیک اسید (DNA) نام دارند. DNA از چهار نوع مولکول ساخته شده است، که به آنها " نوکلئوتید " می گوییم. این چهار نوکلئوتید عبارتند از: ادنین (A)، گوانین (G)، سیتوزین (C) و تیمین (T). این مولکولها خود به دو زیر گروه تقسیم می شوند: ادنین و گوانین در گروه پیورین قرار می گیرند و سیتوزین و تیمین در گروه پیریمیدین جای داده می شوند. هنگام سنتز (ساخته شدن) مولکول DNA، نوکلئوتیدها به اسیدهای نوکلئیک تبدیل می شوند که بعدا به هم متصل می شوند و رشته های DNA را می سازند. در نهایت یک مارپیچ دوتایی ساخته می شود.
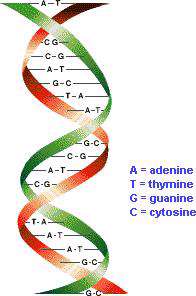
نوکلئوتیدها حلقه های مسطحی دارند که اندازه آنها بین 3 تا 4 آنگستروم ( هر آنگستروم 10-10 متر است.) می باشد. وقتی مارپیچ دوتایی تشکیل می شود، مولکولهای A با مولکولهای T رشته مقابل و مولکول G با مولکول C رشته روبه رو پیوند هیدروژنی برقرار می کنند و جفت های قلیایی ایجاد می شود. این جفت های قلیایی باعث می شوند که مارپیچ پایدار باقی بماند. تصاویری که با استفاده از اشعه X از مولکول DNA گرفته شده است، نشان می دهد که در هر دور از مارپیچ 10 جفت قلیایی وجود دارد.
مدل مارپیچی می تواند نحوه رونویسی از مولکول DNA در هنگام تقسیم سلول را نیز توجیه کند. جیمز واتسون وقتی این مدل را ارائه کرد، آن را " مدل خوش نما " نامید، چون معتقد بود که هر کس این مدل را ببیند می تواند به راحتی نحوه رونویسی از آن را نیز بفهمد. در هنگام تقسیم سلول، باید نسخه مشابهی از DNA تهیه شود تا در سلولهای جدید قرار گیرد. این فرآیند اصطلاحا "رونویسی" نامیده می شوند.
برای انجام این کار، اتصالات هیدروژنی بین نوکلئوتیدها باز می شود و در نتیجه دو رشته مارپیچ از هم باز می شوند. سپس هر یک از این رشته ها به عنوان پایه ای برای ساخت رشته مقابل استفاده می شود. به این ترتیب دو مارپیچ کاملا یکسان DNA ساخته می شوند و هر کدام از آنها در یکی از دو سلول نوزاد قرار می گیرد. چون در هر بار رونویسی، نیمی از مولکول DNA قبلی حفظ می شود، می گوییم رونویسی DNA، نیمه پایستار است.
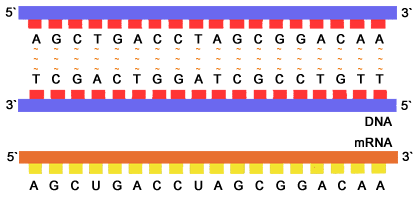
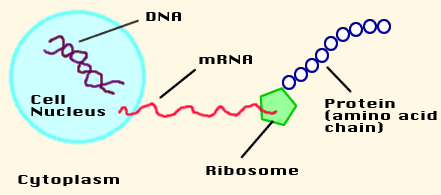
اگرچه DNA اطلاعات ژنتیکی جاندار زنده را در خود دارد، اما برای عملکرد موفق به وجود ریبونوکلئیک اسید (RNA) نیاز دارد. RNA هم مانند DNA از رشته های اسید نوکلئیکی تشکیل شده که با پیوندهای مشابهی به هم متصل شده اند؛ اما دو تفاوت عمده با DNA دارد. یکی اینکه در ساختار آن به جای تیمین، از اوراسیل(u) استفاده شده است و دوم اینکه مارپیچی نیست و فقط از یک رشته تنها ساخته شده است. برای انجام بعضی کارها، DNA به رشته های RNA تبدیل می شود و سپس این مولکولهای RNA، پیغام هایی را به ریبوزوم (مرکز پروتئین سازی سلول) می برند. برای همین این مولکولها را mRNA(messenger RNA) یا RNA پیغام رسان می نامند. در واقع DNA با فرستادن mRNA، فرآیند پروتئین سازی را هدایت می کند.
مقایسه RNA با DNA
RNA و DNA هر دو اسید نوکلئیک هستند، اما در سه چیز تفاوت دارند؛ نخست این که برخلاف DNA که دو رشتهای است، RNA یک مولکول تکرشتهای است و زنجیرهٔ بسیار کوتاه تری از نوکلئوتیدها را دارد. دوم این که در حالی که DNA دارای قند دئوکسیریبوز میباشد، RNA دارای ریبوز است (در دئوکسیریبوز هیچ گروه هیدروکسیلی به حلقه پنتوزی در جایگاه ۲ پیوند ندارد). این گروههای هیدروکسیلی، پایداری RNA را کمتر از پایداری DNA میسازند زیرا داشتن گروه هیدروکسیل ریبوز را برای واکنش آبکافت آمادهتر میسازد. سوم این که برخلاف DNA در RNA، باز تکمیلکنندهٔ آدنین، تیمین نیست بلکه اوراسیل میباشد که شکل متیلینشدهای از تیمین میباشد. بیشتر RNAهای کارا از دیدگاه زیستی که شامل RNA کوچک هستهای، RNA رناتنی، RNA جابجایی، RNA پیامرسان و دیگر RNAهای بیرمز، گه گاه دارای چیدمانهای خود تکمیلکنندهای هستند که به بخش هایی از RNA این اجازه را میدهند که با خودش جفت شده و تا بخورد و مارپیچهای دوتایی را پدید آورند (همانند DNA). برخلاف DNA، ساختار آنها دارای مارپیچهای دوتایی دراز نیستند اما در جای جای آنها گروههایی از مارپیجهای کوتاه دیده میشود.
ساختار
هر نوکلئوتید در آرانای دارای یک قند ریبوز با کربنهای شمارهگذاریشده از ۱ تا ۵ است. یکی از بازهای آدنین، گوانین، سیتوزین، یا اوراسیل به کربن شمارهٔ ۱ پیوند میخورد. به آدنین و گوانین، خانوادهٔ پورینها (دوحلقهایها) گفته میشود و به سیتوزین و اوراسیل، خانوادهٔ پیریمیدینها (تکحلقهایها) گفته میشود. گروههای فسفات دارای یک بار منفی هستند که با پیوند به آرانای، آن را یک مولکول باردار میسازند. بازها ممکن است پیوندهای هیدروژنی میان سیتوزین با گوانین، آدنین با اوراسیل، و گوانین با اوراسیل را تشکیل دهند. به هر حال برهمکنشهای دیگری هم امکانپذیر میباشد، برای نمونه، پیوند یک گروه از بازهای آدنینی به همدیگر در یک برآمدگی یا تترالوپ GNRA که یک جفت باز گوانین–آدنینی دارد. ویژگی ساختاری مهم آرانای که آن را از دیانآ جدا میسازد، داشتن یک گروه هیدروکسیل در کربن شمارهٔ ۲ قند ریبوز است. بودن این گروه به این میانجامد که در شکل هندسی زنجیرهٔ مارپیچی آن با دیانآ تفاوت پیدا کند. دومین نتیجه پیامد داشتن این گروه هیدروکسیل در کربن ۲، در نواحی انعطافپذیری شکلی (تطبیقی) از یک مولکول آرانای است (که در تشکیل یک مارپیچ دوتایی درگیر نیست)، آرانای تنها با چهار باز توصیف میشود که عبارتند از آدنین، سیتوزین، گوانین، و اوراسیل، در آرانای ریبوزومی، بسیاری از اصلاحات پس از رونویسی، در نواحی بسیار عملکردی اتفاق میافتد، از جمله مرکز پپتیدیل ترانسفراز و زیرواحد رابط، که نشاندهندهٔ این است که آنها برای عملکرد عادی، مهم هستند. شکل عملکردی مولکولهای تکرشتهای آرانای، کاملاً مانند پروتئینها، به یک ساختار سوم ویژهای نیاز دارد. چارچوب (داربست) این ساختار توسط عناصر ساختاری دوم تولید میشود که همان پیوندهای هیدروژنی درونمولکولی هستند. این ساختار دوم به پدید آمدن چندین نمایهٔ قابل شناسایی مانند حلقههای سنجاق سری، شکمخوردگیها، و حلقههای درونی میانجامد. از آنجایی که آرانای باردار است، یونهای فلزی از جمله Mg2+ برای پایاسازی بسیاری از ساختارهای دوم و سوم مورد نیاز هستند.
۳۰
دی
واژه کروموزم به مفهوم جسم رنگی ، که در سال 1888 بوسیله والدیر بکار گرفته شد. هم اکنون این واژه برای نامیدن رشتههای رنگپذیر و قابل مشاهده با میکروسکوپهای نوری
بکار میرود که از همانندسازی و نیز بهم پیچیدگی و تابیدگی هر رشته
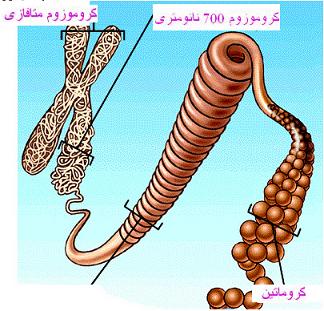
کروماتین اینترفازی در سلولهای یوکاریوتی تا رسیدن به ضخامت 1000 تا 1400
نانومتر ایجاد میشود. در پروکاریوتها نیز ماده ژنتیکی اغلب به حالت یک
کروموزوم متراکم میشود. در برخی باکتریها علاوه بر کروموزوم اصلی که اغلب
ژنها را شامل میشود کروموزوم کوچک دیگری که بطور معمول آن را پلاسمید
مینامند، قابل تشخیص است گر چه تعداد کمی از ژنها بر روی پلاسمید قرار
دارند.
اما از آنجا که در بیشتر موارد ژنهای مقاومت به آنتی بیوتیکها بر روی آن جایگزین شدهاند، از نظر پایداری و بقای نسل باکتری اهمیت زیادی دارد. کروماتین در ساختمان کروموزوم به شکل لوپ دیده میشود. لوپها توسط پروتئینهای اتصالی به DNA که مناطق خاصی از DNA را تشخیص میدهند پابرجا میماند. سپس مراحل پیچ خوردگی نهایتا نوارهایی را که در کروموزومهای متافازی دیده میشود ایجاد میکند. هر تیپ کروموزومی یک نوع نواربندی اختصاصی را در ارتباط با نوع رنگ آمیزی نشان میدهد. این رنگ آمیزیها منجر به مشخص شدن تعداد و خصوصیات کروموزومهای هر گونه از موجودات زنده میگردد. که این خصوصیات تعدادی و مورفولوژیک کروموزومها را کاریوتیپ مینامند.
اولین مرحله پیچیدگی و تراکم رشته کروماتین برای تبدیل به کروموزوم با فسفریلاسیون شدید هیستونهای H3 ، H1 همراه است. پس از رها شدن DNA از اکتامر هیستونی ، با دخالت آنزیمهای مسئول همانندسازی ، پیوندهای هیدروژنی بین دو زنجیره گسسته میشود، هر زنجیره مکممل خود را میسازد و به تدریج با ادامه همانندسازی ، دو مولکول DNA بوجود میآید که در هر مولکول یک زنجیره قدیمی و زنجیره دیگر نوساخت است. بخشهای مختلف این دو مولکول DNA که نظیر همدیگر هستند به تدریج که همانندسازیشان پایان میپذیرد، با اکتامرهای هیستونی که نیمی از آنها اکتامرهای والدی و نیمی جدید هستند ترکیب میشوند.
بعد از تشکیل ساختمان نوکلئوزومی ، دو رشته کروماتین 10 نانومتری و سپس رشتههای 30 نانومتری ایجاد میشوند. هر رشته کروماتین 30 نانومتر سطوح سازمان یافتگی را میگذارند، با مجموعهای از پروتئینهای غیر هیستونی زمینهای یا اسکلتی آمیخته میشود و به یک کروماتید تبدیل میشود. مجموعه دو کروماتید نظیر هم که از محل سانترومر بهم متصلاند کروموزوم متافازی را ایجاد میکنند.
در پروفاز و گاهی در اینترفاز ، کروموزوم به صورت رشتههای بسیار نازکی است که آنها را کرومونما مینامند این رشتهها مراحل مقدماتی تراکم کروماتید را نشان میدهند. کروماتید و کرومونما ، نامی برای مشخص کردن دو ساختمان یکسان اما با دو درجه سازمان یافتگی است. کرومومر نیز از تجمع ماده کروماتینی به صورت دانههای کروی ایجاد میشود.
اما از آنجا که در بیشتر موارد ژنهای مقاومت به آنتی بیوتیکها بر روی آن جایگزین شدهاند، از نظر پایداری و بقای نسل باکتری اهمیت زیادی دارد. کروماتین در ساختمان کروموزوم به شکل لوپ دیده میشود. لوپها توسط پروتئینهای اتصالی به DNA که مناطق خاصی از DNA را تشخیص میدهند پابرجا میماند. سپس مراحل پیچ خوردگی نهایتا نوارهایی را که در کروموزومهای متافازی دیده میشود ایجاد میکند. هر تیپ کروموزومی یک نوع نواربندی اختصاصی را در ارتباط با نوع رنگ آمیزی نشان میدهد. این رنگ آمیزیها منجر به مشخص شدن تعداد و خصوصیات کروموزومهای هر گونه از موجودات زنده میگردد. که این خصوصیات تعدادی و مورفولوژیک کروموزومها را کاریوتیپ مینامند.
|
مراحل تبدیل رشته کروماتین به کروموزوم
برای تبدیل یک رشته کروماتینی 10 تا 30 نانومتری به یک کروموزوم ، علاوه بر لزوم همانندسازی رشته کروماتین سطوح سازمان یافتگیای را در نظر میگیرند که ضمن آن با دخالت H3 ، H1 و پروتئینهای غیر هیستونی پیچیدگیها و تابیدگیهای رشته کروماتین افزایش مییابد، طول آن کم ، ضخامت و تراکمش زیاد میشود و به کروموزوم تبدیل میگردد. این سطوح سازمان یافتگی و اغلب به صورت رسیدن از رشته 10 تا 30 نانومتری به رشته 90 تا 100 نانومتری تشکیل رشته 30 تا 400 نانومتری و در مراحل بعد با افزایش پیچیدگیها و تابیدگیها ، ایجاد رشته 700 نانومتری و بالاخره تشکیل کروموزوم دارای دو کروماتید و با ضخامت تا 1400 نانومتر در نظر میگیرند.اولین مرحله پیچیدگی و تراکم رشته کروماتین برای تبدیل به کروموزوم با فسفریلاسیون شدید هیستونهای H3 ، H1 همراه است. پس از رها شدن DNA از اکتامر هیستونی ، با دخالت آنزیمهای مسئول همانندسازی ، پیوندهای هیدروژنی بین دو زنجیره گسسته میشود، هر زنجیره مکممل خود را میسازد و به تدریج با ادامه همانندسازی ، دو مولکول DNA بوجود میآید که در هر مولکول یک زنجیره قدیمی و زنجیره دیگر نوساخت است. بخشهای مختلف این دو مولکول DNA که نظیر همدیگر هستند به تدریج که همانندسازیشان پایان میپذیرد، با اکتامرهای هیستونی که نیمی از آنها اکتامرهای والدی و نیمی جدید هستند ترکیب میشوند.
بعد از تشکیل ساختمان نوکلئوزومی ، دو رشته کروماتین 10 نانومتری و سپس رشتههای 30 نانومتری ایجاد میشوند. هر رشته کروماتین 30 نانومتر سطوح سازمان یافتگی را میگذارند، با مجموعهای از پروتئینهای غیر هیستونی زمینهای یا اسکلتی آمیخته میشود و به یک کروماتید تبدیل میشود. مجموعه دو کروماتید نظیر هم که از محل سانترومر بهم متصلاند کروموزوم متافازی را ایجاد میکنند.
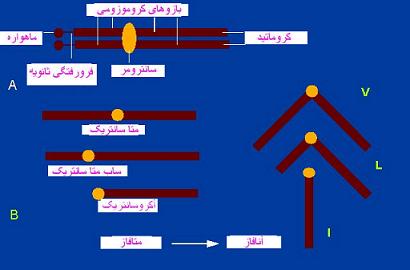
اجزای ساختمانی کروموزوم
در متافاز که کروموزومها سازمان یافتگی بیشتری دارند، برای هر کروموزوم بخشهای زیر در نظر گرفته میشود.کروماتید
کروماتید بخشی از کروموزوم متافازی است که نیمی از سراسر طول کروموزوم را میسازد. دو کروماتید هر کروموزوم از ناحیه سانترومر بهم متصلاند. هر کروماتید از ابر پیچیدگیهای رشته کروماتین و آمیختگی آن با پروتئینهای غیر هیستونی اسکلتی یا زمینهای بوجود آمده است. دو کروماتید هر کروموزوم متافازی را که در حکم تصویر آینهای یکدیگر هستند، کروماتیدهای خواهر یا کروماتیدهای نظیر مینامند.در پروفاز و گاهی در اینترفاز ، کروموزوم به صورت رشتههای بسیار نازکی است که آنها را کرومونما مینامند این رشتهها مراحل مقدماتی تراکم کروماتید را نشان میدهند. کروماتید و کرومونما ، نامی برای مشخص کردن دو ساختمان یکسان اما با دو درجه سازمان یافتگی است. کرومومر نیز از تجمع ماده کروماتینی به صورت دانههای کروی ایجاد میشود.

|
سانترومر
محل اتصال دو کروماتید خواهر هر کروموزوم متافازی را سانترومر نامند. سانترومر بخش نازکی از کروموزوم که جایگاه آنرا فرورفتگی اولیه نیز مینامند. ناحیه سانترومر ناحیه بسیار هتروکروماتینی است و بویژه در بخشهای کناری خود دارای ژنها یا ترتیبهای نوکلوتیدی تکراری است. این بخشهای هتروکروماتین با رنگهای بازی شدت رنگ میگیرند. هر کروموزوم علاوه بر سانترومر اصلی ممکن است دارای سانترومر یا سانترومرهای فرعی در محل فشردگیهای ثانویه باشد. فشردگیهای ثانویه با داشتن پیچیدگیهای کمتر از فشردگی اولیه قابل تشخیصاند.کینه توکور
طرفین سانترمر هر کروموزوم را دو بخش پروتئینی پیاله مانند و متراکم به اسم کینه توکور میپوشاند. هر کینه توکور دارای سه بخش بیرونی و میانی و درونی است. در ساختمان هر بخش پروتئینهای رشتهای با تراکم متفاوتی قابل تشخیص هستند بخش بیرونی متراکم و بخش میانی کم تراکم است. بخش درونی بطور فشردهای با سانترومر اتصال دارد. کینه توکورها از مراکز سازماندهی میکروتوبولها و رشتههای دوک میتوزی هستند.تلومر
این اصطلاح برای بخشهای انتهایی کروماتید بکار گرفته میشود. تلومرها دارای ویژگیهای سلول شناسی خاصی هستند. در مگس سرکه ترتیبهای DNAای تلومری که در انتهای همه کروموزومها وجود دارد جدا سازی و بررسی شده است. تلومرها انتهاهای مولکولهای طویل و خطی DNAای هستند که در هر کروماتید وجود دارد. از سوی دیگر وقتی کروموزومها بوسیله عواملی مثل پرتوهای X یا اثر آلکالوئیدها شکسته شوند، انتهاهای آزاد بدون تلومر آنها چسبنده میشود و با سایر کروموزومها ادغام میشود. علاوه بر نقشی که تلومرها در پایداری کروموزومها دارند، در برخی گونهها به حالت مهیا و بعضی بین دو کروموزوم عمل کرده و نوک به نوک اتصال موقتی پیدا میکنند.فرورفتگی ثانویه
یکی دیگر از ویژگیهای ریخت شناسی کروموزومها هستند که از نظر موقعیت و فواصلشان بر حسب گونهها جای ثابتی دارند. وجود آنها از نظر تشخیص کروموزومها بویژه در یک مجموعه کروموزومی مفید است فرورفتگیهای ثانویه به دلیل عدم ایجاد انحرافهای زاویهدار در قطعات کروموزومی از فرورفتگیهای اولیه شناخته میشوند.سازمان دهندگان هستکی
این نواحی فرورفتگیهای ثانویهای هستند که دارای ژنهای رمزدار کننده RNAهای ریبوزومی جز rRNA5S میباشند و در تشکیل هستک دخالت دارند. پدیدار شدن فرورفتگی ثانویه به دلیل رونویسی بسیار فعال ژنهای rRNAای است که آنها را از فرورفتگیهای اولیه مشخص میسازد. در انسان سازمان دهندگان هستکی در فرورفتگیهای ثانویه کروموزومهای 13 و 14 و 15 و 21 و22 قرار دارند که همه از کروموزمهای آکروسانتریک و دارای ماهواره هستند.ماهواره
جسم کوچکی کروی است که از بقیه کروموزوم بوسیله یک فرورفتگی ثانویه جدا میشود. ماهواره و فرورفتگی ثانویه از نظر شکل و بزرگی برای هر کروموزوم ویژه ، ثابت هستند. ماهوارههای کروموزومی بخشهایی از کروموزوم از دیدگاه ریخت شناسی هستند و نبایستی آنها را با ماهوارههای DNAای که دارای ترتیبهای DNAای بسیار تکراری میباشند اشتباه کرد.
|
انواع کروموزمها از نظر تعداد سانترومر
کروموزومها را از نظر تعداد سانترومرهایشان به کروموزمهای یک سانترومری ، دو سانترومری و چند سانترومری تقسیم میکنند وقتی تحت تاثیر عواملی مثل پرتوهای X کروموزمها خرد شوند و قطعاتشان ادغام شود، کروموزومهای به اصطلاح بدون سانترومر ایجاد میکنند. این کروموزومها هنگام تقسیم سلولی رفتار عادی مثل سایر کروموزومها را ندارند.انواع کروموزوم از نظر محل سانترومر
-
کروموزمهای تلوسانتریک: سانترومر در یکی از دو انتهای کروموزومها قرار گرفته است.
-
کروموزومهای آکروسانتریک:
سانترومر آنها نزدیک به یکی از دو انتهای کروموزوم قرار گرفته در نتیجه
یکی از بازوها نسبتا به دیگری بسیار کوچک است از قطعات کروموزومی از محل
قرار گرفتن سانترومر از بازوهای کروموزومی مینامند.
- کروموزمهای متاسانتریک: سانترومر آنها در وسط کروموزوم قرار گرفته و در نتیجه بازوهای کروموزم هم اندازه هستند اکثر کروموزمها دارای یک سانترومر هستند. برخی گونهها سانترومرهای بخش شدهای دارند در رشتههای دوکی به تمامی طول کروموزوم متصلند این کروموزومها را هولوسانتریک گویند.
۲۲
آذر
فردریک میشر (۱۸۹۵-۱۸۴۴) به سفارش پدرش وارد دانشکدهی پزشکی شد، اما به علت دشواری در شنیدن، نمیتوانست با بیماران به خوبی ارتباط برقرار کند. از این رو تصمیم گرفت، وارد عرصهی پژوهشهای پزشکی شود. وی در سال ۱۸۶۸ پژوهشهای خود را زیر نظر فلیکس هوپ سیلر۶ در دانشکدهی علوم طبیعی دانشگاه توبینگن آلمان آغاز کرد. در آن آزمایشگاه، هنگامی که هنوز بسیاری از دانشمندان در مفهوم «سلول» شک داشتند، برخی از مولکولهای سازندهی سلولها استخراج شده بودند و پژوهش در زمینهی شیمی بافتها ادامه داشت.
بررسی شیمیایی سلولهای سفید خون، به عنوان موضوع پژوهشهای مایشر برگزیده شد. استخراج این سلولها از گرههای لنفاوی بسیار دشوار بود، اما در زخمهای چرک مقدار زیادی از آنها یافت میشود. از این رو، مایشر باندهای آلوده را از بیمارستان محلی جمعآوری و با کمک محلولی از نمک، گلبولهای سفید را از آنها جدا می کرد. مایشر در جریان یکی از آزمایشهایش، گلبولهای سفید را تحت تأثیر عصارهی معدهی خوک قرار داد. در آن زمان، دانشمندان میدانستند این عصاره ، آنزیمی دارد که باعث هضم پروتئینها میشود. امروزه آن آنزیم را با نام پپسین میشناسیم. وی چگونگی اثر عصاره را بر این سلولها، به دقت زیر میکروسکوپ پی گیری کرد. وقتی عصارهی معده ، پروتئینهای سفید خون را تخریب کرد، او مشاهده کرد که ساختار این سلولها از هم پاشید، اما هستهی آنها تا حدود زیادی سالم باقی ماند. به این ترتیب، او هستهی سلولها را از سیتوپلاسم جدا کرد.
در گام بعدی، هستهها را تحت تأثیر هیدروکسید سدیم قرار داد. افزودن این محلول قلیایی به ظرف حاوی هستهها، باعث تشکیل رسوب سفید رنگی شد که تجزیهی شیمیایی آن نشان داد، کربن، هیدروژن، اکسیژن، نیتروژن و درصد زیادی فسفر، عنصر های سازندهی آن هستند. پایداری در برابر عمل پپسین، چگونگی واکنش آن به حلالهای متفاوت و درصد فسفر بالا باعث شد که مایشر پیشنهاد کند، ماده غیر پروتئینی جدیدی را کشف کرده است. وی این ماده را نوکلئین به معنای «در هسته» نامید.
میشر آزمایشهای مشابهی را روی اسپرم ماهی آزاد انجام داد. به طور کلی، هسته در همهی اسپرمها حجم زیادی از سلول را به خود اختصاص میدهد. در اسپرم ماهی آزاد نیز بیش از ۹۰ درصد حجم سلول، از هسته است. تلاش شبانهروزی این پژوهشگر پرکار به استخراج نوکلئین از اسپرم ماهی آزاد و اسپرم گونههای دیگر منجر شد. بررسی شیمیایی نوکلئین استخراج شده از آن منابع، نتیجهی پیشین را تأئید کرد. مایشر بهراستی مادهی جدیدی کشف کرده بود که به نظر میرسید، در هستهی همهی سلولها وجود دارد. آیا این ماده نمیتوانست مادهی ژنتیک باشد؟
اگر نوکلئین مادهی ژنتیک باشد، باید مقدار آن در همهی سلولهای پیکری یکسان و در سلولهای جنسی نصف سلولهای پیکری باشد. مایشر برای بررسی این فرضیه چند سال تلاش کرد و توانست مقدار نوکلئین را در هستهی سلولهای پیکری و جنسی تعیین کرد. اما یک رویداد ناشی از بدشانسی باعث شد، او به اشتباه نوعی پروتئین را به عنوان مادهی ژنتیک معرفی کند.
میشر درصد فسفر بالا را معیار شناسایی نوکلئین قرار داده بود. در سیتوپلاسم سلول تخمک، پروتئینی به نام فسویتین۷ وجود دارد که بر خلاف دیگر پروتئینها، مقدار زیادی فسفر دارد. این پروتئین که در آن زمان کشف نشده بود، باعث شد مایشر مقدار نوکلئین موجود در تخمک را به درستی محاسبه نکند. از این رو، نتیجه گرفته که مقدار نوکلئین سلول تخمک و سلول اسپرم با هم برابر نیستند و بنابراین چنین مولکولی نمیتواند نقش ماد هی ژنتیک را بازی کند.
میشر پس از سالها تلاش، در اثر سل جان باخت. دو عامل را دلیل ابتلای او به این بیماری میدانند: تماس با چرک باندهای بیماران و فعالیت شبانهروزی در اتاق سردی که برای استخراج نوکلئین لازم بود. در هر صورت، وی جان خویش را بر سر شناخت نوکلئین گذاشت.(نامش زنده و یادش گرامی باد)
فوبوس لون (۱۹۴۰-۱۸۶۹) فراگیری پزشکی را در روسیه آغاز کرد، اما به سبب کار در آزمایشگاه شیمی آلی، به زیستشیمی ( بیوشیمی ) علاقه مند شد. در سال ۱۸۲۹ آموزش پزشکی را در نیویورک به پایان رساند و با بزرگان شیمی از جمله آلبرت کوسل۸ و امیل فیشر۹ آشنا شد که در زمینهی اسید نوکلئیک و پروتئین کار می کردند. او در نتیجهی پژوهشهای فراوان ، بیش از ۷۰۰ مقاله دربارهی ساختمان شیمیایی مولکولهای زنده منتشر کرد، اما شهرت او بیشتر به سبب طرح تترانوکلئوتیدی است.
لون براساس پژوهشهای خود و پژوهش گران پیشین به این نتیجه رسید که نوکلئوتیدها واحد ساختمانی اسیدهای نوکلئیک هستند و اسید نوکلئیکی که مایشر کشف کرده بود، از نوع داکسی ریبونوکلئیک (DNA) است. هر نوکلئوتید از یک نوع باز آلی، یک قند پنج کربنه و یک گروه فسفات تشکیل شده که در شرایط طبیعی به صورت یونیزه و دارای بار منفی است. به علاوه او دریافت، نوکلئو تیدها از راه اتصال فسفودی استری به هم پیوند میشوند.
لون براساس آزمایشهای خود به این نتیجهی نادرست دست یافت که اندازهی چهار باز A ، T ، C و G ، در DNA برابر است. از این رو، طرح تترانوکلئوتیدی را به عنوان ساختمان شیمیایی DNA پیشنهاد کرد. براساس این طرح، DNA مولکول درازی است که از تکرار یک واحد تترانوکلئوتیدی (چهار نوکلئوتیدی) تشکیل شده است؛ یعنی، به صورت زیر:
AGTC-AGTC-AGTC-AGTC … )n …)
روشن است که چنین مولکول یکنواختی نمیتوند اطلاعات وراثتی گوناگون جاندارن را در خود اندوخته کند. به این ترتیب، طرح تترانوکلئوتیدی لون از این باور پشتیبانی کرد که با وجود حضور DNA در کروموزومها، این مولکول نمیتواند مادهی وراثتی باشد. البته، این اشتباه نباید نقشی را که لون در شناخت ساختمان شیمیایی DNA داشته است، از یاد ببرد.
اروین چارگاف
اروین چارگاف (۱۹۹۲-۱۹۲۹) در زمینهی شیمی، پژوهشهای گستردهای انجام داده، اما بیش تر به خاطر به دست آوردن نسبت بازهای آلی در DNA مشهور است. وی و همکارانش به مدت هفت سال با روش کروماتوگرافی کاغذی، نسبت بازهای آلی DNA را در جاندارن گوناگون و سلولهای پیکری یک جاندار تعیین کردند و نتیجه گرفتند، مقدار بازها در DNA گونههای مختلف جانداران متفاوت است و با تغییر رژیم غذایی، تغییر شرایط محیطی یا افزایش سن جاندار، تغییر نمیکند. اما در تمام نمونهها، مقدار A با مقدار T و مقدار C با مقدار G برابر است.
آزمایشهای چارگاف نشان داد، نظریهی تترانوکلئوتیدی لون درست نیست. نتیجهی این آزمایشها، در روش ساختن ساختمان مولکولی DNA و چگونگی اندوخته شدن اطلاعات در آن، نقش مهمی داشتند. به هر حال، خود او نتوانست از آنها در این زمینه بهره گیرد.
لینوس پاولینگ
روش پراش پرتوی ایکس نخستین بار برای مطالعهی بلور نمک طعام استفاده شد. شیمیدان بزرگ لینوس پاولینگ، یکی از نخستین کسانی بود که با بهرهگیری از این روش تلاش کرد، ساختمان سه بعدی پروتئینها را روشن کند. وی در مجموعه مقالههایی که در سالهای ۱۹۵۰ و ۱۹۵۱ انتشار داد، مارپیچ آلفا را مهم ترین رکن ساختمان سه بعدی پروتئینها معرفی کرد.
پاولینگ برای DNA نیز طرحی پیشنهاد کرد. در طرح او، DNA از سه رشتهی مارپیچ تشکیل شده بود که بازهای آلی آن در بیرون و ستونهای قند فسفات در درون مولکول قرار داشتند. به علاوه، در طرح او گروههای فسفات به حالت یونیزه و دارای بار منفی نبودند و رشتهها از راه پیوندهای هیدروژنی با هم ارتباط داشتند که بین گروههای فسفات برقرار شده بودند.
براساس آنچه که از شیمی DNA میدانیم، گروههای فسفات همیشه به حالت یونیزه و دارای بار منفی هستند و این معما همچنان باقی است که پاولینگ (برند هی نوبل شیمی) چگونه چنین اشتباهی مرتکب شده است؟ باوجود این، همان طور که در ادامه میآید، شیوهی پژوهشی او تأثیر مهمی بر فعالیت های واستون و کریک داشت.
روزالین فرانکلین
روزالین فرانکلین (۱۹۵۸-۱۹۲۰) در سال ۱۹۵۱ به همراه یکی از دانشجویان به نام رایموند گوسلینگ۱۰، مجموعهای از تصویرهای پراش پرتوی ایکس با کیفیت بالا، از بلور DNA تهیه کرد. او با استفاده از این تصویرها تو انست، ابعاد DNA را محاسبه کند و به درستی نتیجه گرفت که گروههای فسفات در بیرون مولکول DNA قرار دارند. به علاوه تشخیص داد، DNA به دو شکل A و B وجود دارد و شکل راستین DNA ، همان شکل B است. تصویری که او از بلور شکل B تهیه کرد، در روشن شدن ساختمان سه بعدی DNA نقش به سزایی داشت. آن تصویر را موریس ویکلینز (با اجازه یا بدون اجازهی فرانکلین) در اختیار واستون و کریک قرار داده بود.(واتسون در کتاب خود، که با نام مارپیچ مضاعف در ایران منتشر شده است، به این حقیقت اشاره کرده است.)
فرانکلین در سال ۱۹۵۸ در اثر سرطان درگذشت. به نظر میرسد، کار بیش از اندازه با پرتو ایکس در ابتلای او به سرطان مؤثر بوده است.
واستون و کریک
در روزهای پایانی سال ۱۹۵۱، جیمز واتسون (زیستشناس) و فرانسیس کریک (فیزیکدان) با هدف تعیین ساختمان مولکولی DNA ، همکاری خویش را آغاز کردند. آنان میدانستند، مولکول DNA از تعداد زیادی نوکلئوتید تشکیل شده است که به صورت خطی و با کمک اتصالهای فسفودی استری کنار یکدیگر قرار گرفتهاند. از سوی دیگر، در همین سال، پاولینگ مارپیچ آلفا را به عنوان مهمترین رکن ساختمان سه بعدی پروتئینها معرف کرده بود. از این رو، نخستین طرح فرضی برای DNA ، در ذهن این زوج علمی شکل گرفت:
۱٫ DNA رشتهای دراز و مارپیچی شکل از واحدهایی به نام نوکلئوتید است. در این رشته، ستون قند فسفات بسیار منظم و ترتیب بازها بسیار نامنظم است.
وقتی آنان طرح فرضی خود را با ویلکینز در میان گذاشتند، با این پاسخ روبهرو شدند که برای اساس تصویرهای پراش پرتوی ایکس، قطر مولکول DNA بیش از آن است که وجود تنها یک رشته پلینوکلئوتیدی آن را توجیه کند. از این رو، کریک پیشنهاد تازهای را مطرح کرد:
۲٫ مولکول DNA از چند رشتهی پلی نوکلئوتیدی تشکیل شده است که به دور یکدیگر پیچ خوردهاند.
آیا DNA مولکولی دو رشتهای، سه رشتهای یا چهار رشتهای است؟ ارتباط این رشتهها با یکدیگر چگونه است؟ آیا به راستی مولکول DNA ساختمان مارپیچی دارد؟ پاسخ این پرسشها با اطلاعات کمی که در اختیار واتسون و کریک بود، به دست نمیآمد. از این رو، از ویلکینز خواستند با آنان همکاری کند و تصویر پراش پرتوی ایکس بلور DNA را در اختیارشان قرار دهد. آنان با در دست داشتن تصویر پراش پرتوی ایکس DNA ، تصمیم گرفتند همانند دیگر دانشمندانی که به مطالعهی بلور مولکولها میپرداختند، با استفاده از سیم و تکههای حلب، طرح فرضی DNA را بسازند.
تفسیر تصویرهای پراش بلورها، به محاسبهی پیچیدهای نیاز دارد. در آن زمان، هنوز رایانه وارد آزمایشگاههای بلورشناسی نشده بود. از این رو، بلورشناسان با توجه به اطلاعات اندکی که از تصویرهای پراش پرتو ایکس به دست میآوردند، طرحهای فرضی مولکولها را میساختند. سپس با انجام محاسبههایی ، الگوی پراش فرضی این طرحهای ساختگی را تعیین میکردند. سرانجام، پراش فرضی با پراش بلور مقایسه و ساختمان سه بعدی مولکول مورد نظر پیشبینی میشد. برای مثال، وجود تقارن و نظم در تصویر پراش بلور، نشان دهندهی نظم و تکرار واحدهای سازندهی مولکولهای بلور است. بنابراین، طرح ساخته شده باید دارای نظم و واحدهای تکرار شونده باشد.
واتسون و کریک با فرض این که ستون قند فسفات در مرکز و بازهای حلقوی در بیرون مولکول DNA قرار دارند، به ساختن نخستین طرح برای DNA مشغول شوند. براساس این طرح :
۳٫ DNA از دو رشتهی پلی نوکلئوتیدی تشکیل شده است. این رشتهها با پلهای نمکی به هم مربوط میشوند که در آنها کاتیونهای دو ظرفیتی مانند +Mg2 و گروههای فسفات دارای بار منفی، شرکت دارند.
پس از پایان کار، آنان از ویلکینز و فرانکلین دعوت کردند، طرحشان را بررسی کنند. وقتی آنان مسألهی یونهای +Mg2 را مطرح کردند که دو رشته را کنار یکدیگر نگه میدارند، با اعتراض شدید فرانکلین روبهرو شدند. فرانکلین پافشاری کرد که یونهای +Mg2 را پوستههایی از مولکولهای آب دربرمیگیرند و بسیار دور است میخ محکمی برای نگهداشتن ساختمان DNA باشند. نظر او این بود که ستون قند و فسفات در بیرون قرار دارد. به این ترتیب، مولکولهای آب، طرح دو رشتهای واتسون و کریک را فروریختند.
مدتها از این ماجرا گذشت ، بدون آن که واتسون و کریک به موفقیت چشمگیری دست پیدا کنند. تا این که با خبر شدند، پاولینگ برای ساختمان سه بعدی DNA ، طرحی پیشنهاد کرده است. اما همان طور که گفته شد، طرح مارپیچ سه رشتهای پاولینگ از نظر شیمیایی نادرست بود.
مدتی بعد، در دیداری که این زوج علمی با ویکلینز داشتند، با تصویر تازهای از بلور DNA روبهرو شدند که از تصویرهای پیشین سادهتر بود. آن تصویر را که مربوط به شکل B بود، فرانکلین تهیه کرده بود. ویلکینز به آنان گفت، آن تصویر از بلوری تهیه شد ه که مقدار زیادی آب داشته است و تصویر پیشین که آن دو روی آن کار میکردهاند، از مولکولی بوده که آب خود را از دست داده بوده است. کریک به کمک ویلکینز آن تصویر را با معادلههای ریاضی بررسی کرد تا اطلاعات زیر به دست آمد:
۱) تصویر پراش بسیار منظم است. بنابراین، ساختمان مولکولی DNA باید بسیار منظم و قطر آن در همهی مولکول ثابت باشد.
۲) نقش ضربدری که در تصویر مشا هده میشود، از مارپیچ بودن مولکول DNA حکایت میکند و زاویهی بین بازوی ضربدر و خط افق، با زاویهی پیچش DNA برابر است.
۳) در تصویر پراش، نقطههایی که فاصلهی زیادی از هم دارند، در واقع فاصلهی اندکی از یکدیگر دارند و برعکس. با در نظر گرفتن این قاعده که معادلههای پیچیدهی ریاضی آن را تأ یید میکنند، فاصلهی بین مرکز و محیط تصویر پراش، حدود ۳۴ انگستروم و فاصلهی بین هر ردیف از نقطههای سیاه با ردیف بعدی، حدود ۳۴ انگستروم محاسبه میشود. بنابراین، فاصلهی هر جفت باز با جفت باز دیگر، حدود ۴/۳ انگستروم و فاصلهی عمودی یک دور کامل مارپیچ DNA ، حدود ۳۴ انگستروم خواهد بود. در این صورت، در هر دور مارپیچ DNA ، حدود ۱۰ جفت باز آلی جای میگیرد.
سرانجام، واتسون و کریک با درنظر گرفتن این اطلاعات و نتیجهی آزمایشهای چارگاف، توانستند به بزرگترین کشف زیستشناسی مولکولی دست یابند و به همراه ویلکینز، جایز هی نوبل ۱۹۶۲ را از آن خود کنند.
در این میان جای روزالین فرانکلین بسیار خالی بود.
سخن پایانی
کشف مارپیچ دوتایی، نمونهی خوبی از نقش و تأثیر دانشمندان رشتههای گوناگون علوم، در حل یک مسأله است. بدون شناختن ویژگیهای فیزیکی و شیمیایی DNA ، زیستشناسان هرگز نمیتوانستند به این کشف مهم دست پیدا کنند. جالب این که، در این کشف نقش شیمیدانان و فیزیکدانان، از زیستشناسان پررنگتر بود.
فردریک میشر

۲۲
آذر
ساختار DNA و ارتباط آن با نوکلئوتید:
برای درک عملکرد موکول از هر نوعی که باشد ابتدا باید با ساختار آن آشنا شویم.
بلوک های بنیادینDNA به نام نوکلئوتید شناخته میشوند که هر کدام شامل یک قند حلقوی 5 کر بنه به نام دئوکسی ریبوز می باشند.
یک گروه فسفات به صورت استری در موقعیت 5َ قند و یک باز نیتروژنه در موقعیت 1َ آن قرار دارد.
به طور کلی 2 نوع باز در نوکلئیک اسید ها وجود دارد:
1.پورینها : که 2 حلقه ای هستند شامل آدنین و گوانین (A,G)
2.پیریمیدینها : که تک حلقه هستند و شامل سیتوزین و تیمین (C,T)
نوکلئوتیدها به صورت کوالانت به هم متصل می شوند تا یک پلیمر خطی را تشکیل دهند با یک ستون فقرات که از تناوب قند و فسفات تشکیل شده است که با پیوند های فسفو دی استر به هم اتصال دارند .
بازهائی که به هر قند متصلند همانند طبقات روی هم به نظر می رسند
یک نوکلئوتید دارای ساختار قطبی است: یک انتها که فسفات قرار گرفته انتهای 5َ نامیده می شود و انتهای دیگر آن 3َ است
در ابتدا تصور میشد زنجیره DNA از توالی ثابت 4 باز آلی A , T , C , G تشکیل شده است اگر توالی DNA به این صورت باشد در همه گونه ها هر نوع باز باید 25 درصد از کل ماده ژنتیک را تشکیل دهد در صورتی که آزمایشات چارگف نشان داد نسبت 4 باز در جانداران مختلف با هم تفاوت عمده ای دارد .
و قانون چارگف به این صورت بیان شد که تعداد پورین ها و پیریمیدین ها در یک DNA با هم برابر است واین روابط بین بازهای آلی وجود دارد:
A=T , C=G , A+T≠ C+G
کشف DNA


اهداف:
- آشنایی با ساختار DNA و طرز کشف آن
چطور دانشمندان متوجه شدند که DNA ساختاری دو رشته و به هم تاب خورده است؟
چطور دانشمندان متوجه شدند که بازه ای چهار گانه به این ترتیب که ادنین در مقابل تیمین و سیتوزین در مقابل گوانین قرار دارد؟
برای پاسخ به این سوالات باید کمی به قبل برگردیم. زمانی که دستگاه های پیشرفته وجود نداشت و دانشمندان به طور غیر مستقیم به این ساختار رسیدند.
اروین چارگف
در سال 1949 با جداسازی DNA از 4 گونه مختلف و اندازه گیری هر یک از4 باز
موجود پی برد که میزان باز ادنین به باز تیمین بسیار نزدیک است و میزان باز
گوانین و سیتوزین نیز تقریباً مساوی است. در همین زمان پاولینگ و همکارش
با استفاده از تکنیک کریستالوگرافی اشعه ایکس پی به ساختار مارپیچی آلفا
هلیکس در پروتئین ها بردند. وقتی اشعه ایکس به DNA تابیده شود برخی از اشعه
ها پس از بازگشت و پراکندگی از مولکول DNA به فیلم فوتوگرافیک برخورد کرده
و ساختار DNA را روی فیلم به طور سایه نشان می دهد. از روی همین فیلم
دانشمندان توانستند اندازه هر مارپیچ را متوجه شوند.
اروین چارگف پی برد که میزان باز ادنین به باز تیمین بسیار نزدیک است و میزان باز گوانین و سیتوزین نیز تقریباً مساوی است. وقتی اشعه ایکس به DNA تابیده شود برخی از اشعه ها پس از بازگشت و پراکندگی از مولکول DNA به فیلم فوتوگرافیک برخورد کرده و ساختار DNA را روی فیلم بطور سایه نشان می دهد. از روی همین فیلم دانشمندان توانستند اندازه هر مارپیچ را متوجه شوند. همچنین این گروه با کنار هم قرار دادن بازهای مختلف و اندازه گیری طول هر جفت باز به رابطه صحیح بین باز ها با هم پی بردند.
هم چنین این دانشمندان با توجه به تجارب خود از بیوشیمی و بارهای موجود در گروه های قند و فسفات آن ها را در کنار هم قرار داند. هم چنین چون ساختار مارپیچ DNA به طور یکنواخت است انها با کنار هم قرار دادن بازهای مختلف و اندازه گیری طول هر جفت باز به رابطه صحیح بین باز ها با هم پی بردند.
در اخر نیز واتسون و کریک با کمک همدیگر و با استفاده از تجارب دانشمندان قبلی مدل فعلی DNA را معرفی کردند.
اما چرا باید DNA به طور مارپیچی باشد و چه نفعی برای سلول دارد که DNA به این شکل باشد؟
فرم مارپیچی DNA سبب کاهش تغییرات آنتالپی می گردد و هر فرم دیگر به غیر از این سبب هدر رفتن انرژی می گردد.
۲۲
آذر

این وبلاگ اخبار و اتفاقات مدرسه علامه حلی ۱ از زبان آن ها (مخصوصا دوره۳۲)نشر میدهد
طبقه بندی موضوعی
-
تکالیف
(۱۲) -
علمی وجذاب
(۱۲) -
اخبار مدرسه
(۲۴) -
کامپیوتر
(۸) -
متون
(۸)-
انجمن فارسی سره
(۱)
-
-
انشا
(۲) -
اجتماعی
(۱۳) -
زیست شناسی
(۲۲) -
کار وفناوری
(۷) -
دینی
(۷)-
قرآن
(۱)
-
-
زبان
(۱) -
عربی
(۴) -
فیزیک
(۹) -
ریاضی و هندسه
(۱۲) -
شیمی
(۸) -
اخبار سایت
(۷) -
پویش
(۲)
بایگانی
- ارديبهشت ۱۳۹۵(۱)
- فروردين ۱۳۹۵(۱)
- اسفند ۱۳۹۴(۳)
- بهمن ۱۳۹۴(۱۸)
- دی ۱۳۹۴(۳۹)
- آذر ۱۳۹۴(۶۶)
- آبان ۱۳۹۴(۳۵)
آخرین مطلب
-
۹۵/۰۲/۲۵زیست
-
۹۵/۰۱/۲۲زیست
-
۹۴/۱۲/۱۲while
-
۹۴/۱۲/۱۲فردا آزمونIMC
-
۹۴/۱۲/۱۲مندلیف
-
۹۴/۱۱/۱۷برنامه جدید کلاس 1/3
-
۹۴/۱۱/۱۶الن تورینگ
-
۹۴/۱۱/۱۶تمرین ۹ کامپیوتر ۱/۳
-
۹۴/۱۱/۱۶تکلیف پویش ۲
-
۹۴/۱۱/۱۶آموزش تبدیل word به pdf
پربیننده ترین مطالب
-
۹۴/۱۱/۰۷جذر
-
۹۴/۰۹/۱۷آلوتروپی های کربن(مقایسه ساختار الماس و گرافیت)
-
۹۴/۱۰/۰۷نمونه سوال درس ۱۸ اجتماعی با جواب
-
۹۴/۰۹/۱۲دانلود کتاب شیمی دان شکاک(به زبان انگلیسی)
-
۹۴/۰۹/۱۸استان شناسی خوزستان
-
۹۴/۱۱/۰۵ویدیو های زیست (رونویسی و ترجمه)
-
۹۴/۱۰/۰۸جزوه
-
۹۴/۰۹/۱۲طرز کار ترازو ۳ کفه ای
-
۹۴/۰۸/۲۷پاور پوینت ایران شناسی(اصفهان)(به مناسبت اردو)
-
۹۴/۰۸/۲۷با وبلاگ آشنا شوید!
محبوب ترین مطالب
-
۹۴/۰۸/۲۷با وبلاگ آشنا شوید!
-
۹۴/۰۹/۲۲جدول چگالی
-
۹۴/۰۸/۲۰سمینار علوم و فنون چیست؟
-
۹۴/۰۹/۱۱آمار
-
۹۴/۰۸/۱۷جواب امتحان هندسه
-
۹۴/۰۹/۱۰زیست
-
۹۴/۰۹/۱۲طرز کار ترازو ۳ کفه ای
-
۹۴/۰۹/۱۲جواب امتیازی شیمی (آمونیوم دی کرومات)
-
۹۴/۰۸/۱۷لیست بچه ها هفتم
-
۹۴/۰۸/۱۹نمونه سوال عربی
مطالب پربحثتر
-
۹۴/۱۰/۰۸جزوه
-
۹۴/۰۸/۲۷با وبلاگ آشنا شوید!
-
۹۴/۰۹/۱۸استان شناسی خوزستان
-
۹۴/۱۰/۲۰نمونه سوال کامپیوتر(طرح شده توسط آقای صالحه)
-
۹۴/۱۰/۱۳نمونه سوال دینی (برای ۲۰ گرفتن فقط کافیست حلی ۳۲ ای شوید)
-
۹۴/۰۸/۲۷یک ایموجی کلمه ی سال شد!
-
۹۴/۰۹/۱۲جواب امتیازی شیمی (آمونیوم دی کرومات)
-
۹۴/۰۹/۱۸تمرین شماره 8 ریاضی
-
۹۴/۱۰/۰۶مصاحبه با معلمان مدرسه
-
۹۴/۱۰/۲۲جزوه زیست
نویسندگان
- علی توفیقی(98)
- بهراد صادقی(37)
- آرین کسایی(28)